Living cell membranes are made of phospholipids assembled in twodimensional bilayers in such a way that the hydrophobic chains are being shielded from the surrounding water as illustrated in Figure 1. Due to their extreme softness the phospholipid bilayers exhibit thermal undulations in an aqueous environment that give rise to repulsive forces between the cells or between cells and substrates. These forces govern important phenomena in the life process such as cell coagulation and the ability of the cells to sense and react to their surroundings. Here, we report results on the thermal undulations of phospholipid bilayers obtained through the Neutron Spin Echo ( NSE ) method, the only technique covering the time scale of these motions, 1 ns to 10 ns [ 1, 2 ].
The thermal undulations of phospholipid bilayers are strongly dependent on the mechanical properties of the bilayer such as the bending elasticity. Following earlier studies by other methods [ 3, 4 ] we have studied the dynamics of a model system, multilamellar vesicles ( MLV, see Figure 1 ) in water, as a function of temperature, the presence of cholesterol, and the presence of salt. Temperature is expected to influence several properties. Increase in temperature decreases the solvent viscosity and increases the thermal undulation amplitudes. The hydrophobic core of the phospholipid membranes exhibits a sharp phase transition from the crystalline to the liquid phase at a certain temperature Tc that depends on the length of the hydrophobic chains. Above Tc the interior of the phospholipid membrane is much more disordered and is expected to be less rigid. Cholesterol, an important constituent of the living cell, nestles in between the fatty tails, changing the mechanical properties of the bilayers and hence their motions. Finally, the presence of sodium chloride as an electrolyte has a screening effect on electrostatic interactions and may affect the thermal undulations of electrostatically charged bilayers.
Three MLV samples are prepared for this study. Sample 1 ( “vesicles” ), which serves as a reference, is a MLV made mainly of 1,2-Dimyristoyl-sn-Glycero-3- Phosphocholine ( DMPC ) in deuterated water ( D2O ) to provide scattering contrast. The DMPC vesicles were stabilized through enhanced electrostatic repulsion between the bilayers by addition of a small amount ( 0.05 mol to 1 mol DMPC ) of 1,2-Dimyristoyl-sn-Glycero-3- [ Phosphorac- ( 1-glycerol ) ] -sodium salt ( DMPG ), a phospholipid that is structurally similar to DMPC. In sample II ( “vesicles & chol” ) cholesterol was added in the ratio of 0.3 mol cholesterol to 1 mol of lipids. In sample III ( “vesicles & salt” ) 0.05 mol L-1 of N a C l was added. MLV were prepared by extrusion through a filter with a 200 nm pore size that predetermines their average diameter, as confirmed by dynamic light scattering.
The NSE technique provides a measurement of the real component of the intermediate scattering function in the time domain I( Q,t ). Thermal diffusive motions lead to an exponential time decay of I ( Q,t ). Therefore we have fitted our data for each sample at each temperature with
, where the effective diffusion coefficient De f f is a sum of two parts: Dd e f, representing the diffusive motions associated with the fluctuations within each vesicle, the thermal undulations; and Dtr, the translational diffusion coefficient of the vesicles in the solution [ 5 ] . The latter can be neglected because the vesicles prepared here are quite large, R
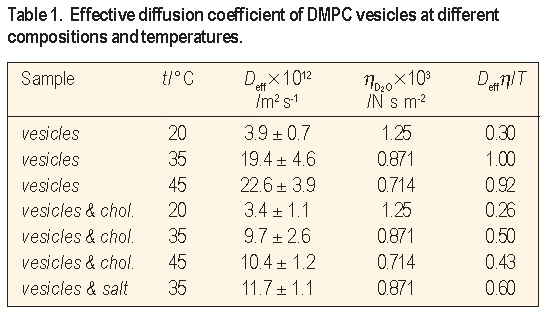
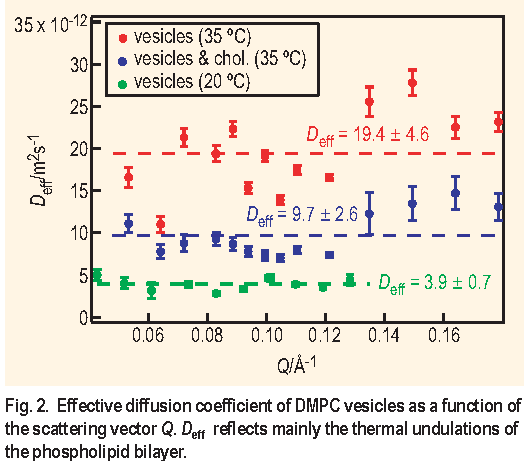
100 nm, and according to the Stokes-Einstein hydrodynamic equation the corresponding translational diffusion Dtr is less than 2.6 X 10-12 m2 s-1. Samples "vesicles" and "vesicles & chol." were studied at temperatures of 20 °C, 35 °C and 45 °C to cover regions below and above Tc of DMPC which is 24 °C. Sample "vesicles & salt" was measured at 35 °C to study the effect of added electrolyte. Selected results are shown on Figure 2 and summarized in Table 1.
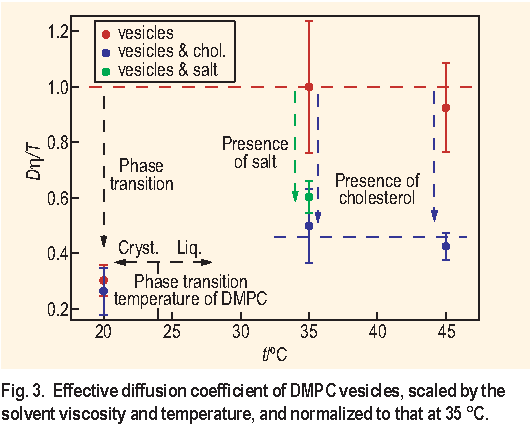
To take into account the effect of the solvent viscosity and temperature we have calculated the ratio De f f
/T and normalized it to that at 35 °C. All deviations of the normalized parameter De f f
The analysis of the normalized parameter De f f
- Scaling the diffusion coefficient De f f that measures the thermal undulations of DMPC bilayers like De f f
- The bilayer liquid to crystalline phase transition increases the rigidity of DMPC bilayers by a factor of 3 for vesicles and by a factor of 1.5 for vesicles in presence of cholesterol.
- Cholesterol increases the rigidity of the DMPC bilayers in a bilayer-liquid state by a factor of 2 but has no effect on the DMPC bilayers in a bilayer-crystalline state.
- Addition of 0.05 mol L-1 of N a C l reduces the fluctuations of the DMPC bilayers by a factor of 2. This effect can be attributed either to the reduced electrostatic repulsion between the DMPC bilayers or to the effect of the osmotic pressure that can deflate the MLV.
These results are in good agreement with those obtained from other methods [ 3, 4 ] . They are a direct confirmation that cholesterol stiffens cell membranes thus reducing the cell — cell repulsions that originate from undulation forces. The significant difference in the thermal undulations below and above the bilayer liquid to crystalline phase transition temperature shows the importance of temperature for the proper functioning of the cell membranes. Finally, it has been shown that the NSE method provides a direct means of observing thermal undulation of biological membranes.
References:
[1] W. Pfeiffer, S. Köning, J. F. Legrand, T. Bayerl, D. Richter, and E. Sackmann, Europhys. Lett. 23, 457 (1993).
[2] T. Takeda, Y. Kawabata, H. Seto, S. Komura, S. K. Shosh, M. Nagao, and D. Okuhara, J. Phys. Chem. Solids 60, 1375 (1999).
[3] H. P. Duwe, and E. Sackmann, Physica A 163, 410 (1990).
[4] C. Hofsäß, E. Lindahl, and O. Edholm, Biophys. J. 84, 2192 (2003).
[5] S. T. Milner, and S. A. Safran, Phys. Rev. A 36, 4371 (1987).
D. P. Bossev and N. S. Rosov
NIST Center for Neutron Research
National Institute of Standards and Technology
Gaithersburg, MD 20899-8562
Back to FY2003 HTML main page
Go to previous article
Go to next article
To view all symbols correctly, please download Internet Explorer 6 or Netscape 7.1